В 40-60-х годах нашего столетия К. Блох и сотр. в опытах с использованием ацетата, меченного 14С по метильной и карбоксильной группам, показали, что оба атома углерода уксусной кислоты включаются в холестерин печени приблизительно в одинаковых количествах. Кроме того, было доказано, что все атомы углерода холестерина происходят из ацетата.
В дальнейшем благодаря работам Ф. Линена, Г. Попьяка, Дж. Корн-форта, А.Н.
Климова и других исследователей были выяснены основные детали ферментативного синтеза холестерина, насчитывающего более 35 энзиматических реакций.
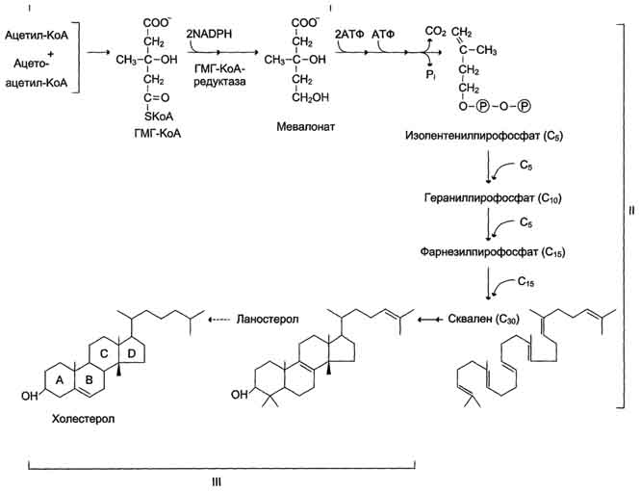
В синтезе холестерина можно выделить три основные стадии: I – превращение активного ацетата в мевалоновую кислоту, II – образование сквалена из мевалоновой кислоты, III – циклизация сквалена в холестерин.
Рассмотрим стадию превращения активного ацетата в мевалоновую кислоту. Начальным этапом синтеза мевалоновой кислоты из ацетил-КоА является образование ацетоацетил-КоА посредством обратимой тиолазной реакции:
Затем при последующей конденсации ацетоацетил-КоА с 3-й молекулой ацетил-КоА при участии гидроксиметилглутарил-КоА-синтазы (ГМГ-КоА-синтаза) образуется β-гидрокси-β-метилглутарил-КоА:
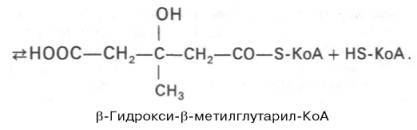
Далее β-гидрокси-β-метилглутарил-КоА под действием регуляторного фермента НАДФ-зависимой гидроксиметилглутарил-КоА-редуктазы (ГМГ-КоА-редуктаза) в результате восстановления одной из карбоксильных групп и отщепления HS-KoA превращается в мевалоновую кислоту:
ГМГ-КоА-редуктазная реакция – первая практически необратимая реакция в цепи биосинтеза холестерина. Она протекает со значительной потерей свободной энергии (около 33,6 кДж). Установлено, что данная реакция лимитирует скорость биосинтеза холестерина.
Наряду с классическим путем биосинтеза мевалоновой кислоты имеется второй путь, в котором в качестве промежуточного субстрата, по-видимому, образуется не β-гидрокси-β-метилглутарил-КоА, а β-гидрокси-β-метилглутарил-S-АПБ.
Реакции этого пути идентичны начальным стадиям биосинтеза жирных кислот вплоть до образования ацетоацетил-S-АПБ. В образовании мевалоновой кислоты по этому пути принимает участие ацетил-КоА-карбоксилаза – фермент, осуществляющий превращение ацетил-КоА в малонил-КоА.
Оптимальное соотношение малонил-КоА и ацетил-КоА для синтеза мевалоновой кислоты – 2 молекулы ацетил-КоА на 1 молекулу малонил-КоА.
Участие малонил-КоА – основного субстрата биосинтеза жирных кислот в образовании мевалоновой кислоты и различных полиизопреноидов показано для ряда биологических объектов: печени голубя и крысы, молочной железы кролика, бесклеточных дрожжевых экстрактов.
Этот путь биосинтеза мевалоновой кислоты отмечен преимущественно в цитозоле клеток печени. Существенную роль в образовании мевалоната в данном случае играет ГМГ-КоА-редуктаза, обнаруженная в растворимой фракции печени крысы и неидентичная микросомному ферменту по ряду кинетических и регуляторных свойств.
Регуляция второго пути биосинтеза мевалоновой кислоты при ряде воздействий (голодание, кормление холестерином, введение поверхностно-активного вещества тритона WR-1339) отличается от регуляции первого пути, в котором принимает участие микросомная редуктаза.
Эти данные свидетельствуют о существовании двух автономных систем биосинтеза мевалоновой кислоты. Физиологическая роль второго пути окончательно не изучена.
Полагают, что он имеет определенное значение не только для синтеза веществ нестероидной природы, таких, как боковая цепь убихинона и уникального основания N6-(Δ2-изопентил)-аденозина некоторых тРНК, но и для биосинтеза стероидов (А.Н. Климов, Э.Д. Полякова).
На II стадии синтеза холестерина мевалоновая кислота превращается в сквален. Реакции II стадии начинаются с фосфорилирования мевалоновой кислоты с помощью АТФ. В результате образуется 5-фосфорный эфир, а затем 5-пирофосфорный эфир мевалоновой кислоты:
5-пирофосфомевалоновая кислота в результате последующего фосфорилирования третичной гидроксильной группы образует нестабильный промежуточный продукт – 3-фосфо-5-пирофосфомевалоновую кислоту, которая, декарбоксилируясь и теряя остаток фосфорной кислоты, превращается в изопентенилпирофосфат. Последний изомеризуется в диметилаллилпирофосфат:
Затем оба изомерных изопентенилпирофосфата (диметилаллилпирофос-фат и изопентенилпирофосфат) конденсируются с высвобождением пи-рофосфата и образованием геранилпирофосфата:
К геранилпирофосфату вновь присоединяется изопентенилпирофосфат. В результате этой реакции образуется фарнезилпирофосфат:
В заключительной реакции данной стадии в результате НАДФН-за-висимой восстановительной конденсации 2 молекул фарнезилпирофосфата образуется сквален:
На III стадии биосинтеза холестерина сквален под влиянием сквален-оксидоциклазы циклизируется с образованием ланостерина.
Дальнейший процесс превращения ланостерина в холестерин включает ряд реакций, сопровождающихся удалением трех метильных групп, насыщением двойной связи в боковой цепи и перемещением двойной связи в кольце В из положения 8, 9 в положение 5, 6 (детально эти последние реакции еще не изучены):
Приводим общую схему синтеза холестерина:
Начиная со сквалена, все промежуточные продукты биосинтеза холестерина (включая и холестерин) нерастворимы в водной среде. Поэтому они участвуют в конечных реакциях биосинтеза холестерина, будучи связанными со стеринпереносящими белками (СПБ). Это обеспечивает их растворимость в цитозоле клетки и протекание соответствующих реакций.
Данный факт имеет важное значение и для вхождения холестерина в клеточные мембраны, окисления в желчные кислоты, превращения в стероидные гормоны. Как отмечалось, реакцией, регулирующей скорость биосинтеза холестерина в целом, является восстановление β-гидрокси-β-метилглутарил-КоА в мевалоновую кислоту, катализируемое ГМГ-КоА-редуктазой.
Данный фермент испытывает регуляторное воздействие ряда
факторов. В частности, скорость синтеза редуктазы в печени подвержена четким суточным колебаниям: максимум ее приходится на полночь, а минимум – на утренние часы.
Активность ГМГ-редуктазы возрастает при введении инсулина и тире-оидных гормонов. Это приводит к усилению синтеза холестерина и повышению его уровня в крови.
- При голодании, тиреоидэктомии, введение глюкагона и глюкокорти-коидов, напротив, отмечается угнетение синтеза холестерина, что прежде всего связано со снижением активности ГМГ-КоА-редуктазы.
- Предыдущая страница | Следующая страница
- СОДЕРЖАНИЕ
- Еще по теме:
- Биосинтез холестерина – Наглядная биохимия
Источник: http://www.xumuk.ru/biologhim/169.html
Холестерол. Его химическое строение, биосинтез и биологическая роль
Стероиды – производные восстановленных конденсированных циклических систем – циклопентанпергидрофенантренов.
В организме человека основной стероид – холестерол, остальные стероиды – его производные.
Растения, грибы и дрожжи не синтезируют холестерол, но образуют разнообразные фитостеролы и микостеролы, не усваиваемые организмом человека. Бактерии не способны синтезировать стероиды.
Холестерол входит в состав мембран и влияет на структуру бислоя, увеличивая её жёсткость. Из холестерола синтезируются жёлчные кислоты, стероидные гормоны и витамин D3. Нарушение обмена холестерола приводит к развитию атеросклероза.
Холестерол представляет собой молекулу, содержащую 4 конденсированных кольца, обозначаемые латинскими буквами А, В, С, D, разветвлённую боковую цепь из 8 углеродных атомов в положении 17, 2 “ангулярные” метальные группы (18 и 19) и гидроксильную группу в положении 3. Наличие гидроксильной группы позволяет относить холестерол к спиртам, поэтому его правильное химическое название “холестерол”, однако в медицинской литературе часто используют термин “холестерин”.
Присоединение жирных кислот сложноэфирной связью к гидроксильной группе приводит к образованию эфиров холестерола.
В неэтерифицированной форме холестерол входит в состав мембран различных клеток. Гидроксильная группа холестерола обращена к водному слою, а жёсткая гидрофобная часть молекулы погружена во внутренний гидрофобный слой мембраны.
В крови 2/3 холестерола находится в этерифицированной форме и 1/3 – в виде свободного холестерола. Эфиры холестерола служат формой его депонирования в некоторых клетках (например, печени, коры надпочечников, половых желёз). Из этих депо холестерол используется для синтеза жёлчных кислот и стероидных гормонов.
Синтезируется во многих тканях человека, но основное место синтеза – печень. В печени синтезируется более 50% холестерола, в тонком кишечнике – 15- 20%, остальной холестерол синтезируется в коже, коре надпочечников, половых железах. В сутки в организме синтезируется около 1 г холестерола; с пищей поступает 300-500 мг .
Холестерол выполняет много функций: входит в состав всех мембран клеток и влияет на их свойства, служит исходным субстратом в синтезе жёлчных кислот и стероидных гормонов. Предшественники в метаболическом пути синтеза холестерола превращаются также в убихинон – компонент дыхательной цепи и долихол, участвующий в синтезе гликопротеинов.
Сложный путь синтеза холестерола можно разделить на 3 этапа.
Первый этап заканчивается образованием мевалоната (мевалоновой кислоты). Две молекулы ацетил-КоА конденсируются ферментом тиолазой с образованием ацетоацетил-КоА.
Фермент щдроксиметилглутарил-КоА-синтаза присоединяет третий ацетильный остаток с образованием ГМГ-КоА (3-гидрокси-3-метилглутарил-КоА). Эта последовательность реакций сходна с начальными стадиями синтеза кетоновых тел. Однако реакции синтеза кетоновых тел происходят в митохондриях печени, а реакции синтеза холестерола – в цитозоле клеток.
Следующая реакция, катализируемая ГМГ-КоА-редуктазой, является регуляторной в метаболическом пути синтеза холестерола. В этой реакции происходит восстановление ГМГ-КоА до мевалоната с использованием 2 молекул NADPH. Фермент ГМГ-КоА-редуктаза – гликопротеин, пронизывающий мембрану ЭР, активный центр которого выступает в цитозоль.
На втором этапе (Образование сквалена) синтеза мевалонат превращается в пятиуглеродную изопреноидную структуру, содержащую пирофосфат – изопентенилпирофосфат. Продукт конденсации 2 изопреновых единиц – геранилпирофосфат.
Присоединение ещё 1 изопреновой единицы приводит к образованию фарнезилпирофосфата – соединения, состоящего из 15 углеродных атомов.
Две молекулы фарнезилпирофосфата конденсируются с образованием сквалена – углеводорода линейной структуры, состоящего из 30 углеродных атомов.
На третьем этапе (Образование холестерола) синтеза холестерола сквален через стадию образования эпоксида ферментом циклазой превращается в молекулу ланостерола, содержащую 4 конденсированных цикла и 30 атомов углерода. Далее происходит 20 последовательных реакций, превращающих ланостерол в холестерол. На последних этапах синтеза от ланостерола отделяется 3 атома углерода, поэтому холестерол содержит 27 углеродных атомов.

Образование кетоновых тел в тканях. Биологическое значение кетоновых тел. Кетоацидоз.
При голодании, длительной физической работе и в случаях, когда клетки не получают достаточного количества глюкозы, жирные кислоты используются многими тканями как основной источник энергии.
В отличие от других тканей мозг и другие отделы нервной ткани практически не используют жирные кислоты в качестве источника энергии.
В печени часть жирных кислот превращается в кетоновые тела, которые окисляются мозгом, нервной тканью, мышцами, обеспечивая достаточное количество энергии для синтеза АТФ и уменьшая потребление глюкозы. К кетоновым телам относят β-гидроксибутират, ацетоацетат и ацетон.
Первые две молекулы могут окисляться в тканях, обеспечивая синтез АТФ. Ацетон образуется только при высоких концентрациях кетоновых тел в крови и, выделяясь с мочой, выдыхаемым воздухом и потом, позволяет организму избавляться от избытка кетоновых тел.
Синтез кетоновых тел в печени. При низком соотношении инсулин/глюкагон в крови в жировой ткани активируется распад жиров. Жирные кислоты поступают в печень в большем количестве, чем в норме, поэтому увеличивается скорость β-окисления.
Скорость реакций ЦТК в этих условиях снижена, так как оксалоацетат используется для глюконеогенеза. В результате скорость образования ацетил-КоА превышает способность ЦТК окислять его. Ацетил-КоА накапливается в митохондриях печени и используется для синтеза кетоновых тел.
Синтез кетоновых тел происходит только в митохондриях печени.
Ацетоацетат может выделяться в кровь или превращаться в печени в другое кетоновое тело – β-гидроксибутират путём восстановления.
В клетках печени при активном β-окислении создаётся высокая концентрация NADH. Это способствует превращению большей части ацетоацетата в β-гидроксибутират, поэтому основное кетоновое тело в крови – именно β-гидроксибутират. При голодании для многих тканей жирные кислоты и кетоновые тела становятся основными топливными молекулами.
Глюкоза используется в первую очередь нервной тканью и эритроцитами. При высокой концентрации ацетоацетата часть его неферментативно декарбоксилируется, превращаясь в ацетон. Ацетон не утилизируется тканями, но выделяется с выдыхаемым воздухом и мочой.
Таким путём организм удаляет избыточное количество кетоновых тел, которые не успевают окисляться, но, являясь водорастворимыми кислотами, вызывают ацидоз.
Окисление кетоновых тел в периферических тканях
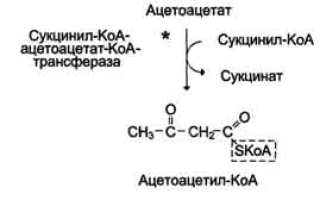
β-Гидроксибутират, попадая в клетки, дегидрируется NAD-зависимой дегидрогеназой и превращается в ацетоацетат. Ацетоацетат активируется, взаимодействуя с сук-цинил-КоА – донором КоА:
Ацетоацетат + Сукцинил-КоА → Ацетоацетил- КоА + Сукцинат.
Реакцию катализирует сукцинил-КоА-ацето-ацетат-КоА-трансфераза. Этот фермент не синтезируется в печени, поэтому печень не использует кетоновые тела как источники энергии, а производит их “на экспорт”.
Кетоновые тела – хорошие топливные молекулы; окисление одной молекулы β-гидроксибутирата до СО2 и Н2О обеспечивает синтез 27 молекул АТФ.
Эквивалент одной макроэргической связи АТФ (в молекуле сукцинил-КоА) используется на активацию ацетоацетата, поэтому суммарный выход АТФ при окислении одной молекулы β-гидроксибутирата – 26 молекул.
Кетоацидоз. В норме концентрация кетоновых тел в крови составляет 1-3 мг/дл (до 0,2 мМ/л), но при голодании значительно увеличивается. Увеличение концентрации кетоновых тел в крови называют кетонемией, выделение кетоновых тел с мочой – кетонурией.
Накопление кетоновых тел в организме приводит к кетоацидозу: уменьшению щелочного резерва (компенсированному ацидозу), а в тяжёлых случаях – к сдвигу рН (некомпенсированному ацидозу), так как кетоновые тела (кроме ацетона) являются водорастворимыми органическими кислотами (рК~3,5), способными к диссоциации: СН3-СО-СН2-СООН ↔ СН3-СО-СН2-СОО- + Н+.
Ацидоз достигает опасных величин при сахарном диабете, так как концентрация кетоновых тел при этом заболевании может доходить до 400-500 мг/дл. Тяжёлая форма ацидоза – одна из основных причин смерти при сахарном диабете. Накопление протонов в крови нарушает связывание кислорода гемоглобином, влияет на ионизацию функциональных групп белков, нарушая их кон-формацию и функцию.
Источник: https://infopedia.su/16×7221.html
Формула и процесс биосинтеза холестерина. Как рассчитать холестерин низкой плотности зная ЛПВП и общий холестерин
Холестерин относится к группе стероидов. Является одним из главных стероидов в макроорганизме человека, определяет активность обмена липидов. По своей структуре это твердое кристаллическое бесцветное вещество, не растворяющееся в воде. Лабораторной единицей измерения в периферической крови является ммоль/л.
- Химическая формула (она же брутто-формула) холестерина — C27H46O.
- Молекулярная масса — около 387 г/моль.
- Структурная форма выглядит следующим образом:
- Структурная формула холестерола с нумерацией атомов в молекуле
Одна из основных особенностей молекулы холестерола – способность связываться с другими соединениями, образовывая комплексы молекул. Такими соединениями могут быть кислоты, амины, протеины, холекальциферол (предшественник витамина Д3), соли и прочие. Данное свойство обусловлено характерным строением молекулы холестерола и его высокой активностью в процессах биохимии.
Весь холестерин в человеческом макроорганизме подразделяется на экзогенный и эндогенный. Экзогенный составляет около 20% от общего показателя и поступает в организм с продуктами питания. Эндогенный холестерол синтезируется непосредственно в организме. Его производство синхронно происходит в двух локализациях.
В кишечнике специфическими клетками энтероцитами формируется около 15% вещества, а порядка 50% эндогенного холестерина вырабатывается в печени, где в дальнейшем связывается с белками, образует комплексы в виде липопротеидов и выходит в периферический кровоток.
Небольшая часть также отправляется на синтез триглицеридов – эфиров жирных кислот и глицерина, которые соединяются с холестеролом.
Синтез холестерола – сложный и энергозатратный процесс. Необходимо больше 30 последовательных реакций липидной трансформации, чтобы в результате образовалась холестериновая молекула. Схематически, все эти превращения можно сгруппировать в шесть стадий процесса синтеза холестерола.
- Биосинтез мевалоната. Состоит из трех реакций. Первые две из них являются реакциями кетогенеза, а третью реакцию катализирует фермент ГМГ-SКоА редуктаза, под действие которой образуется первый предшественник холестерина – мевалоновая кислота. Механизм действия большинства гиполипидемических препаратов, в особенности статинов, направлен именно на это звено биосинтеза. Путем воздействие на ферментативную активность редуктаз, можно частично управлять холестериновой трансформацией.
- Биосинтез изопентенилпирофосфата. Три фосфатных остатка присоединяются к полученной мевалоновой кислоте. После этого она проходит процессы декарбоксилирования и дегидрирования.
- На третьем этапе происходит слияние трех изопентенилпирофосфатов, которые превращаются в фарнезилдифосфат.
- Из 2-х остатков фарнезилдифосфата образуется новая молекула – сквален.
- Линейный сквален проходит ряд реакций циклизации и трансформируется в ланостерол.
- От ланостерина отщепляются избыточные метильные группы, соединение проходит ступень изомеризации и восстановления, в результате которых образуется молекула холестерина.
Кроме активного фермента ГМГ-КоА редуктазы, в реакциях биосинтеза принимают участие инсулин, глюкагон, адреналин и специальный белок-переносчик, который связывает метаболиты на разных этапах.
При каких показателях следует начинать беспокоиться?
В целом, норма холестерина в крови должна варьироваться от 3 до 6 миллимоль на 1 литр крови. Рассмотрим, что означает, если с помощью анализа на общий холестерин выяснилось, что этот показатель больше или меньше нормы.
Значения ниже нормы могут указывать на следующее:
- Острые заболевания.
- Вирусные инфекции.
- Человек голодает.
- Хронические заболевания легких.
- Болезни суставов.
- Умственные отклонения.
- Человек принимает препараты, которые сжигают холестерин.
- Человек находится на диете и пытается снизить уровень холестерола в крови с помощью употребления в пищу продуктов с большим содержанием полиненасыщенных аминокислот.
Значения, превышающие норму, могут указывать на следующее:
- Болезни печени.
- Почечную недостаточность.
- Может быть симптомом возникновения сахарного диабета.
- Может указывать на опухоль простаты или поджелудочной.
- У женщин повышенный холестерин наблюдается во время беременности.
- При алкоголизме уровень данного жироподобного соединения в крови также возрастает.
- Употребление в пищу продуктов, которые увеличивают уровень холестерола в крови.
- Может быть симптомом ишемической болезни сердца.
- Подагру.
В большинстве случаев, данный анализ назначают для того, чтобы выяснить наличие атеросклероза или ишемической болезни сердца.
Кроме этого, рекомендуют сдавать липидограмму хотя бы раз в год, для предупреждения возникновения различных заболеваний сердечнососудистой системы и болезней печени.
Отклонение в норме показателей в любую сторону – это плохой признак, но в любом случае, точный диагноз способен распознать и назвать только врач и только после проведенного осмотра.
Определение уровня холестерина
Уровень сахараМужчинаЖенщинаУкажите Ваш сахар или выберите пол для получения рекомендацийУровень0.58Идет поискНе найденоУкажите возраст мужчиныВозраст45Идет поискНе найденоУкажите возраст женщиныВозраст45Идет поискНе найдено
Липидограмма (тест на уровень холестерина) назначается при сердечно-сосудистых патологиях, повышении артериального давления, подозрении на диабет, печеночной или почечной дисфункции, а также при проведении скрининговых обследований. К такомуисследованию необходимо правильно подготовиться.
За 9-12 часов до сдачи анализа нельзя ничего есть или пить (речь идет о кофе, чае, алкоголе и прочих напитках). В этот период можно пить только воду. В случае приема медикаментозных средств данный вопрос нужно обсудить с врачом, ведь они могут повлиять на результаты исследования.
Рекомендуется сдавать анализ на холестерин у своего семейного врача. Специалист уже знает семейный анамнез, артериальное давление, сопутствующие патологии и индивидуальные особенности организма. При положительном результате он сможет разработать эффективную схему лечения.
Рассчитывают холестерин по таким показателям, как ЛПВП, ЛПНП и триглицериды. Для этого производится забор капиллярной крови натощак. Норма холестерола в крови в зависимости от пола пациента представлена в таблице.
| Пол | Коэффициент общего холестерина, ммоль/л | ЛПВП, ммоль/л | ЛПНП, ммоль/л |
| Женский | 3,61-5,25 | 0,91-1,91 | 3-4 |
| Мужской | 3,61-5,25 | 0,71-1,71 | 2,24-4,81 |
Даже высокая концентрация общего холестерина не всегда говорит о каких-либо патологиях. Она может быть обусловлена повышенным содержанием ЛПВП. Интересен тот факт, что у некоторых женщин уровень ЛПВП может повышаться в результате воздействия эстрогена.
Коэффициент холестеринового отношения подсчитывают следующим образом: общий холестерин делят на показатель ЛПВП. Например, общее содержание холестерола равно 10 ммоль/л, а уровень ЛПВП – 2 ммоль/л, тогда холестериновое отношение составляет 5:1.
- https://holestein.ru/analizy/formula-i-sintez-holesterina
- http://DlyaSerdca.ru/xolesterin/obshhij-xolesterin-v-krovi.html
- https://diabetik.guru/info/raschet-holesterina.html
Источник: https://pro-acne.ru/diabet/sintez-holesterina-formula-strukturnaya-himicheskaya-i-biohimiya-protsessa.html
Формула и процесс биосинтеза холестерина. Общий холестерин и его фракции
Холестерин относится к группе стероидов. Является одним из главных стероидов в макроорганизме человека, определяет активность обмена липидов. По своей структуре это твердое кристаллическое бесцветное вещество, не растворяющееся в воде. Лабораторной единицей измерения в периферической крови является ммоль/л.
- Химическая формула (она же брутто-формула) холестерина — C27H46O.
- Молекулярная масса — около 387 г/моль.
- Структурная форма выглядит следующим образом:
- Структурная формула холестерола с нумерацией атомов в молекуле
Одна из основных особенностей молекулы холестерола – способность связываться с другими соединениями, образовывая комплексы молекул. Такими соединениями могут быть кислоты, амины, протеины, холекальциферол (предшественник витамина Д3), соли и прочие. Данное свойство обусловлено характерным строением молекулы холестерола и его высокой активностью в процессах биохимии.
Весь холестерин в человеческом макроорганизме подразделяется на экзогенный и эндогенный. Экзогенный составляет около 20% от общего показателя и поступает в организм с продуктами питания. Эндогенный холестерол синтезируется непосредственно в организме. Его производство синхронно происходит в двух локализациях.
В кишечнике специфическими клетками энтероцитами формируется около 15% вещества, а порядка 50% эндогенного холестерина вырабатывается в печени, где в дальнейшем связывается с белками, образует комплексы в виде липопротеидов и выходит в периферический кровоток.
Небольшая часть также отправляется на синтез триглицеридов – эфиров жирных кислот и глицерина, которые соединяются с холестеролом.
Синтез холестерола – сложный и энергозатратный процесс. Необходимо больше 30 последовательных реакций липидной трансформации, чтобы в результате образовалась холестериновая молекула. Схематически, все эти превращения можно сгруппировать в шесть стадий процесса синтеза холестерола.
- Биосинтез мевалоната. Состоит из трех реакций. Первые две из них являются реакциями кетогенеза, а третью реакцию катализирует фермент ГМГ-SКоА редуктаза, под действие которой образуется первый предшественник холестерина – мевалоновая кислота. Механизм действия большинства гиполипидемических препаратов, в особенности статинов, направлен именно на это звено биосинтеза. Путем воздействие на ферментативную активность редуктаз, можно частично управлять холестериновой трансформацией.
- Биосинтез изопентенилпирофосфата. Три фосфатных остатка присоединяются к полученной мевалоновой кислоте. После этого она проходит процессы декарбоксилирования и дегидрирования.
- На третьем этапе происходит слияние трех изопентенилпирофосфатов, которые превращаются в фарнезилдифосфат.
- Из 2-х остатков фарнезилдифосфата образуется новая молекула – сквален.
- Линейный сквален проходит ряд реакций циклизации и трансформируется в ланостерол.
- От ланостерина отщепляются избыточные метильные группы, соединение проходит ступень изомеризации и восстановления, в результате которых образуется молекула холестерина.
Пирог с яблоками без сахара и глютена простой рецепт
Кроме активного фермента ГМГ-КоА редуктазы, в реакциях биосинтеза принимают участие инсулин, глюкагон, адреналин и специальный белок-переносчик, который связывает метаболиты на разных этапах.
Холинэстераза (ХЭ)
Норма в сыворотке крови: 5300 — 12900 Ед/л (160-340 ммоль/ч*л).Различают два типа холинэстераз: так называемую истинную холинэстеразу (ацетилхолинэстераза, найденная вэритроцитах, нервной и мышечной ткани) и псевдохолинэстеразу (сывороточная холинэстераза).
Истинная ХЭ катализирует распад ацетилхолина до холина и уксусной кислоты, а псевдоХЭ — распад бутирилхолина.
Сывороточная холинэстераза секретируется печенью, содержится в сыворотке крови, печени, поджелудочной железе, выводится из организма путем фильтрации через почки.
Причины повышения, уровня холинэстеразы в сыворотке крови:
- Причины повышенной холинэстеразы:
- артериальная гипертония
- нефроз
- рак молочной железы
- ожирение
- алкоголизм
- сахарный диабет
- столбняк
- маниакально-депрессивный психоз
- Холинэстераза бывает понижена на фоне:
- заболеваний печени (цирроз, гепатит, метастатический рак печени)
- острого отравления фосфорорганическими соединениями
- инфаркта миокарда
- онкологических заболеваний
Основной причиной снижения активности холинэстеразы (ХЭ) в сыворотке крови служит нарушение белоксинтезирующей функции печени.
Так, уменьшение активности холинэстеразы на 30-50% характерно для острого гепатита и длительно текущего хронического гепатита, а на 50-70% — для циррозов и опухолей с метастазами в печень.
Однако чаще всего уровень холинэстеразы в сыворотке определяется для диагностики отравлений фосфорорганическими инсектицидами, а также для выявления атипичных форм фермента у больных перед хирургическими операциями для предотвращения эффекта рекураризации на фоне применения миорелаксантов.
Рекураризация — повторное угнетение нервно-мышечного синапса после восстановления его функции к концу операции, клинически проявляющееся повторной миорелаксацией и угнетением дыхания. Одним из мероприятий по предупреждению этого грозного осложнения является своевременная
декураризация
путем
введения
антихолинэстеразных препаратов.
Определение концентрации свободного и этерифицированного холестерина в сыворотке крови
Свободный холестерин способен образовывать с дигитонином, томатином, пиридинсульфатом труднорастворимые соединения. Чаще всего применяют водно‑спиртовый или изопропаноловый раствор дигитонина.
Полезные свойства маточного молочка при сахарном диабете
Принцип
Холестерин экстрагируют из сыворотки с помощью изопропилового спирта, экстракт делят на две части, в одной определяют содержание общего холестерина.
В другой порции экстракта свободный холестерин осаждают дигитонином, супернатант отбрасывают, а осадок растворяют и определяют содержание свободного холестерина любым методом.
Содержание этерифицированного холестерина рассчитывают как разницу между общим и свободным.
Нормальные величины
| Сыворотка | фракция этерифицированного холестерина составляет 60‑80% от общего |
Клинико‑диагностическое значение
Коэффициент этерификации холестерина является важной функциональной пробой печени. Снижение коэффициента пропорционально снижению функции печени: острый и обострения хронического гепатита, механическая желтуха, цирроз печени.
Степень этерификации также зависит от активности сывороточного фермента лецитин-холестерин-ацил-трансферазы, поэтому хранение пробы при комнатной температуре может изменить соотношение между свободной и этерифицированной фракцией холестерина.
Лизоцим
Лизоцим (мурамидаза) — антибактериальный агент, вырабатываемый макрофагами, моноцитами и нейтрофилами. Лизоцим гидролизует полисахариды клеточной стенки граммположитедьных бактерий и разрушает их клеточные оболочки.
Фермент содержится в местах соприкосновения организма с окружающей средой: в слезной жидкости, слюне, слизи носоглотки, в слизистой оболочке желудочно-кишечного тракта и т. д., а также в моче и сыворотке крови и придает биологическим жидкостям антибактериальные свойства.
Благодаря малому размеру молекул лизоцим проходит через почечный фильтр и реабсорбируется в почечных канальцах.
- Лизоцим (норма):
- в сыворотке крови — 8-12 мг/л
- моче — 1,3 — 3,6 мг/сут
- слюне — 1,7±0,2 мг/л
- слезной жидкости — 0,96 — 1,44 г/л
Определение активности имеет определенное диагностическое значение при гематологических (лейкоз, гранулематоз), нефрологических, инфекционных и эндокринных заболеваниях.Причины повышенного лизоцима в сыворотке:— моноцитарные и миелоцитарные лейкозы;— полицитемия;— заболевания почек с нарушением фильтрации;— туберкулез;— саркоидоз.
Уровень лизоцима снижается в сыворотке при нейтропении. Содержание фермента в моче увеличивается при повреждении проксимальных канальцев почек, а в слюне — при синдроме Шегрена (аутоиммунное повреждение слюнных и слезных желез).
6 лучших глюкометров – рейтинг 2018 (ТОП-6)
Определение содержания общего холестерина ферментативным методом по набору “Новохол”
Принцип
Основан на использовании сопряженных ферментативных реакций, катализируемых: 1) холестеролэстеразой, катализируещей гидролиз эфиров холестерина до свободного холестерина; 2) холестеролоксидазой, катализирующей превращение холестерина в холестенон с образованием перекиси водорода; 3) пероксидазой, катализирующей в присутствии фенола окисление перекисью водорода 4‑аминоантипирина с образованием окрашенного продукта розово‑малинового цвета.
Нормальные величины
| Сыворотка(указанный метод) | 20‑29 лет | 3,70‑6,51 ммоль/л |
| 30‑39 лет | 4,25‑7,04 ммоль/л | |
| 40‑49 лет | 4,37‑7,70 ммоль/л | |
| старше 50 лет | 4,55‑8,24 ммоль/л | |
| Спинномозговая жидкость | 0‑0,013 ммоль/л | |
| Слюна | 0,065‑0,233 ммоль/л | |
| Желчь | печеночная | 2,1‑5,4 ммоль/л |
| пузырная | в среднем 11,1 ммоль/л |
Влияющие факторы
Завышение результатов при колориметрических методах исследования происходит при высоком содержании в пробе билирубина, гемоглобина, витамина A; при ферментативном методе — оксикортикостероидов и применении антикоагулянтов (фторидов, оксалатов).
Клинико‑диагностическое значение
Сыворотка
Существенное повышение содержания холестерина отмечается при гиперлипопротеинемии IIa типа (семейная гиперхолестеринемия), IIb и III типа (полигенная гиперхолестеринемия, семейная комбинированная гиперлипидемия), умеренное повышение наблюдается при гиперлипопротеинемии I, IV, V типа, а также заболеваниях печени (внутри‑ и внепеченочный холестаз), заболеваниях почек, злокачественных опухолях поджелудочной железы, гипотиреозе, заболеваниях сердечно‑сосудистой системы, беременности, сахарном диабете.
Снижение выявляется при гипертиреозе, циррозе печени, злокачественных опухолях печени, гипопротеинемии и аb‑липопротеинемии.
Спинномозговая жидкость
Накопление холестерина выявляется при менингите, опухоли или абсцессе мозга, кровоизлияниях в мозг, при рассеяном склерозе.
Снижение значений обнаруживается при церебральной и кортикальной атрофии.
Определение количества общего холестерина в сыворотке крови методом Златкис‑Зака
Принцип
Свободный и эфирносвязанный холестерин окисляется хлорным железом в присутствии уксусной, серной и фосфорной кислот с образованием ненасыщенных продуктов, окрашенных в фиолето‑красный цвет.
Нормальные величины
| Сыворотка (указанный метод) | 3,1‑6,5 ммоль/л |
- https://holestein.ru/analizy/formula-i-sintez-holesterina
- https://infolibrum.ru/analysis/ferments/plazmospetsificheskie-fermenty-krovi.html
- https://biokhimija.ru/lipidny-obmen/cholesterin.html
Источник: https://stopzaraza.com/diabet/sintez-xolesterina-formula-strukturnaya-ximicheskaya-i-bioximiya-processa.html
ПОИСК
Стерины. Наиболее важен холестерин (полный синтез его осуществил Р. Вудворд), содержащийся во всех животных клетках (особенно в клетках нервных тканей). Организм человека содержит примерно 140 г холестерина. По строению к холестерину близок растительный стерин — эргостерин. [c.401]
Полный синтез холестерина [c.703]
Р. Б, Вудворд сообщил о полных синтезах холестерина и кортизона. Последний был получен с очень малым вы-хо-дом, но в чистом виде —как один из 64 возможных стереоизомеров. [c.687]
Следующий этап истории полного синтеза, который приходится на 50-е годы, был связан с развитием новых стереохимических представлений, с разработкой специфических реакций, обеспечивших возможность направленного формирования центров асимметрии, и, наконец, с резким увеличением интереса к полному синтезу стероидов, вызванным широким внедрением их в медицинскую практику. В этот период, начавшийся в 1951 г., когда группы Робинсона (схемы 56, 57, 59) и Вудворда (схемы 109, 110) опубликовали работы о полном синтезе андростерона, холестерина (8) и кортизона (6), был осуществлен полный синтез всех неароматических стероидов. В свете сказанного выше о трудности таких синтезов становится [c.10]
Переход от частичного синтеза к полному потребовал методов синтеза гидрированных полициклических соединений так как такие методы отсутствовали, Робинсон [88] (1933 г.) предпринял серию блестящих синтетических изысканий, которые впервые привели к полному синтезу холестерина (см. ниже). 5 [c.232]
Р. Вудворд завершил полный синтез холестерина и кортизона. [c.607]
В последние годы удалось осуществить полный синтез ряда стероидов, в частности холестерина.
Трудность синтеза отдельных стероидов становится очевидной, если учесть, что в случае холес1-ернна, имеющего 8 асимметрических атомов, теоретически возможно 2 = 256 стереонзо-меров.
У описанного ниже эквиленина, обладающего двумя центрами асимметрии, могут существовать только 4 изомера поэтому не случайно, что именно этот стероид был синтезирован первым (см. стр. 873—876). [c.865]
С. выделяют из спинного мозга и желчи рогатого скота, из щелочного гидролизата дрожжей, растит, масел и животных жиров, отходов целлюлозно-бумажВ ой пром-сти, синтезируют из неприродиого сырья. Полный синтез осуществлен для мн. природных С.-холестерина, андрогенов, гестагенов, кортикостероидов, эстрогенов и др. [c.437]
ПОЛНЫЙ СИНТЕЗ СТЕРОИДОВ. СИНТЕЗЫ ЭКВИЛЕНИНА, ЭСТРОНА, ПРОГЕСТЕРОНА, ХОЛЕСТЕРИНА. СИНТЕЗ ЭПИАНДРОСТЕРОНА ПО РОБИНСОНУ [c.394]
Следует подчеркнуть, что Вудворд не был единственным ученым, успешно осуществившим полный синтез стероидов. Одновременно с ним Робинсон опубликовал свой синтез холестерина, правда, гораздо более сложный, а через год последовали синтезы кортикоидов, осуще-стпленныр Сареттом (США). Робинсон исходил из доступного 1,6-диок-синафталина — скелета системы колец В и С. [c.406]
Полный синтез природного холекальциферола (витамина D3) через холестерин формально можно считать осуществленным после того, как в 1951 — 1952 гг. Вудварт с сотр. [163, 164] из простых веществ синтезировал природный холестерин. Синтез холекальциферола из холестерина через 7-дегидрохолестерин и его фотоизомеризацию (с. 132) к тому времени был уже известен. [c.120]
Первый пример полного синтеза стероидов был описан в 1939 г. Бахманном, осуществившим вместе с сотрудниками линейную синтетическую цепочку, при которой последовательно проводилось построение одного за другим колец А, В, С и О. Примером подобного синтеза может служить и синтез холестерина по Вудворду [3.7.3].
В последнее время ( ыли разработаны конвергентные сходящиеся) синтетические цепочки при этом сначала получают два фрагмента целевой молекулы по раздельно осуществляемым схемам, а затем, по возможности на одной из последних стадий, соединяют эти фрагменты в более сложную структуру.
Такой путь синтеза позволяет уменьшить потери промежуточно синтезируемых соединений, на получение которых ступень за ступенью затрачивается много времени и средств. Поэтому конвергентные пути синтеза со сходящимися цепочками особенно привлекательны для промышленного производства стероидов.
Ниже приводится пример построения стероидного скелета с помощью линейного синтеза из метилового эфира 5-оксогептен-6-овой-1 кислоты и 2-метилциклопентандиона-1,3 [3.7.4] (см. схему на с. 696). [c.695]
Ключевой продукт 1 был далее стандартными трансформациями превращен в эфир 20, послуживщий общим исходным соединением для синтеза природных стероидов 2-7.
Селективное гидрирование двух сопряженных двойных связей над палладием привело к продукту 21, в котором сохранив-щаяся изолированная двойная связь в цикле В обеспечивала в дальнейшем возможность введения кислородного заместителя в положение 11. Этот путь позволил осуществить успешный синтез кортикостероидов, в том числе кортизона (7).
Для получения стероидов 2—6 потребовалось полное гидрирование всех трех двойных связей С=С над платиновым катализатором, приводящее к насыщенному эфиру 22. Кроме удаления лишних функциональных групп, эта реакция приводила к созданию требуемой конфигурации хиральных центров С-10 и С-17.
Поскольку превращение эфира 22 в стероиды 2—5 уже было описано ранее, выполненный Вудвордом синтез этого соединения представлял собой одновременно и завершение полного синтеза стероидов 2-5. Наконец, с помощью ряда обьршых методов (присоединение алифатического заместителя в положение 17 и трансформации функциональных групп) из эфира 22 был синтезирован холестерин (6). [c.293]
Холестерин открыт М. Э. Шеврёлем в 1815 г. Как установлено в настоящее время, биосинтез холестерина проходит теми же основными путями, что и биосинтез тритерпенов (см. с. 693).
Установление строения холестерина потребовало свыше столетия работы многих поколений химиков и было закончено лишь в 1934 г.
окончательно подтверждено химическим синтезом холестерина, явившимся одним из первых полных синтезов веществ стероидной природы, осуществленным Р, Б. Вудвордом в 1951 г. [c.704]
Холестерин был открыт еще в начале XIX в. Более ста лет потребовалось для установления его строения (1934). Полный синтез холестери- на, начиная от моноцнклнчсского соединения (метил-/г-.
инона), который включал около 50 сгадн , был осуществлен выдающимся американским. чимнком-органиком Р. Б. Вудвордом (1951).
При полной практической бесполезности в чисто утилитарном смысле такой синтез продемонстрировал могущество человеческого разума. [c.440]
Реакции Дильса — Альдера для хинонов представляют собой один из наиболее интенсивно изучавшихся процессов [26, 39].
В пятидесятые годы несколько лабораторий показали, что региО и стереоселективность этих реакций циклоприсоединения можно использовать в синтезе природных соединений в результате мнО гие полные синтезы (например, холестерина, кортизона, эстрона,, резерпина, террамицина и иохимбина) включают на ранних ста-диях циклоприсоединение к хинону. В полном синтезе холестерина (31), предложенном Вудвордом [схема (14)], циклоприсоеди-нение хинона (32) к бутадиену давало единственный изомер с ожидаемым г ыс-сочленением циклов, который можно изомеризовать в гранс-изомер действием основания [40]. Это приводило к правильной стереохимии сочленения колец С и О в конечном продукте— холестерине (31). [c.842]
Стероиды представляют интерес для химика-органика с начала 1930-х годов, когда впервые была предложена правильная структура холестерина.
Сначала это были попытки получить столь важные молекулы в лабораторных условиях в качестве проверки искусства синтеза и для подтверждения структуры, установленной главным образом путем деградации.
Позднее, по мере возрастания значения многих стероидов как лекарственных препаратов, стало необходимым пополнить количество этих веществ, выделяемое из природного сырья.
Для решения этой последней задачи нет необходимости исходить из простых материалов, как в случае полного синтеза можно использовать имеющиеся в изобилии природные стероиды (например, сапогенины, полученные из растительных гликозидов), которые содержат многие из желаемых структурных признаков и могут быть превращены в ва жные для медицины прогестерон и кортизон в результате небольшого числа стадий с высокими выходами. [c.314]
Результаты, полученные при дегидрировании холестерина, долгое врем не признавались достоверным доказательством его строения, так как продут дегидрирования удается выделить лишь с ничтожным выходом и трудно бы. решить, какой из них образовался без перегруппировки.
Более убедительнь доказательства строения стероидного ядра были получены при окислительнс расщеплении холестерина на многоосновные кислоты с более простым строение скелета. Окончательно зтот вопрос был решен, когда Вудвордом (1951 г.) был ос% ществлен полный синтез холестанола по следующей схеме.
[c.174]
Помимо природных с., в настоящее время известно очень большое число синтетических С. . Последние могут представлять собой продукты как полного, так и частичного синтеза на базе природного стероидного сырья.
Полный синтез осуществлен и для многих природных с. (холестерин, андрогены, эстрогены, гестогены, кортикостероиды и т. д.). Среди синтетических с. выделяется по своей значимости группа 19-норстероидов (см. Норстероиды).
[c.532]
Полные синтезы стероидных соединений осуществлены, но представляют значительные трудности для промышленной реализации.
Обычно используют частичные синтезы из предшественников, содержащих стероидный скелет.
Такими предшественниками являются холестерин (185) и соласодин (188), из которых получают дегидроэпиандростерон (186) его ацетат (187) и ацетат дегидропрегненолона (189) [c.118]
В настоящее время в промышленном масштабе осуществляется лишь полный синтез эстрона, который образуется из дешевого исходного продукта — неролипа — в 8 стадий с общим выходом при учете последних усовершенствований 27% (схемы 38, 39).
Этот метод удовлетворяет всем условиям, необходимым для промышленного полного синтеза и, возможно, выдержит конкуренцию со стороны разработанного в 1965 г. комбинированного химико-микробиологического метода получения эстрона из холестерина [22].
[c.14]
Для построения кольца В необходимо было превратить дикетон (15) в Д -Зр-оксипроизводное (20). С этой целью в -энантиомер соединения (15) путем бромирования и последующего дегидробромирования была введена Д -связь и получен непредельный дикетон (18).
Образование из последнего енолацетата и последовательное восстановление его калием в жидком аммиаке и алюмогидридом лития привело к диолу (19).
3-Окси-группа в нем защищалась трифенилметильной группировкой, 14-окси-групна окислялась по Оппенауэру и после гидролиза защитной группировки был получен кетон (20) [629, 633], идентичный так называемому кетону Кестера — Логеманна, образующемуся при окислении холестерина [634].
В дальнейших реакциях использовалось соединение (20), полученное из природных источников, и поэтому синтез Робинсона следует считать формальным полным синтезом. [c.180]
При использовании других методов введения ангулярного заместителя и сужения кольца D из i u -A/B-оксикетона (162) были осуществлены полные синтезы конессина, прогестерона и холестерина, описанные в предварительных сообщениях (схема 100). [c.248]
В то время как взаимосвязь сквалена и холестерина уже была установлена, только в 1955 г.
[10] удалось показать, что ланостерин может быть синтезирован из уксусной кислоты под действием ферментов, входящих в состав гомогенатов печени крыс, и далее может превращаться в холестерин с потерей трех атомов углерода.
Дальнейшие исследования обнаружили тот замечательный факт, что сквален, меченный С , может превращаться разныл1и способами в зависимости от системы ферментов. Системами, осуществляющими полный синтез холестерина [c.417]
Синтезы более полно гидрированных стероидов, в том числе холестерина, были в последние годы выполнены различными исследователями Ч Мищером и Аннером, Вудвордом, Робинсоном и Корнфорсом, Джонсоном, Сареттом, Веттстепном. Частично эти синтезы были осуществлены в производственном масштабе (Монсанто, Мерк, Циба и др.) [c.865]
После завершения опытов и удаленна трития камеру для разряда откры вали и твердые продукты растворяли в спирте. Аликвоты этих растворов анализировали жидкостным сцинтилляцнопным счетчиком на определенне общего количества введенного трития.
Тритий определяли но потере активности после отгонки спирта. Пальмитиновую кислоту очищали рекристаллизацией из ацетона методом хроматографии по Норнту, а во втором опыте — путем превращения ее в эфир и-бромистого фенацнла.
Бензойная кислота очищалась до постоянной активности путем неоднократной рекристаллизации из воды и гексана. Исходные растворы бензойной кислоты и холестерина обесцвечивали углем. Холестерин частично очищали неоднократной рекристаллизацией из спирта и гептана.
Полная очистка холестерина достигалась путем синтеза его в дибролшд и рекристаллизации этого продукта из смеси эфира и спирта. [c.93]
ПОЛНОГО торможения образования холестерина или резких сдвигов в его обмене.
Следует учитывать, что холестерин является жизненно важным биохимическим компонентом организма, необходимым для нормальной деятельности клеток он является исходным продуктом для синтеза целого ряда необходимых для организма веществ желчных кислот, витамина О, гормонов коры надпочечников, половых гормонов. Препарат трипаранол [c.124]
Семейная гиперхолестеринемия [1087 4440] (рис. 4.71). Семейная гиперхолестеринемия может быть обусловлена приблизительно десятком различных мутаций одного локуса в 19-й хромосоме, которые влияют на работу рецепторов LDL.
Эти мутации подразделяются на несколько классов 1) мутации, нарушающие синтез рецепторов 2) мутации, нарушающие транспорт синтезированного рецептора на клеточную поверхность 3) мутации, нарушающие связывание с LDL 4) мутации, препятствующие ком-партментализации 5) мутации, нарушающие образование кластеров рецепторов в окаймленных пузырьках. Любая из этих мутаций может вызвать тот или иной дефект или полное отсутствие рецепторной функции. Примерно 1/500 часть людей гетерозиготна по наследственной гиперхолестеринемии. У таких индивидов нормальна лишь половина LDL-рецепторов, поэтому обычная скорость удаления холестерина из кровотока не достигается. Это приводит к развитию атеросклероза и сердечным приступам в сравнительно молодом возрасте. Около 50% мужчин-гетерозигот к 50 годам приобретают явные признаки ишемической болезни (разд. 3.8.14). Оказалось, однако, что у гетерозигот можно стимулировать работу нормального аллеля и добиться увеличения синтеза рецепторов LDL, вводя секвестранты желчи (например, холестерин-амин), удаляющие из кишечника желчные кислоты [593]. Этот терапевтический прием, совместно с лечением мевинолином (аналог субстрата HMG СоА-редуктазы) [722], блокирующим синтез холестерина, позволяет понизить уровень холестерина в крови и таким образом воспрепятствовать развитию ишемической болезни. Секвестранты желчи используются уже много лет и зарекомендовали себя как вполне безопасные препараты. [c.122]
У гомозигот с дефектным рецептором вследствие очень высокого содержания липидов рано развивается ишемическая болезнь, и, как правило, они умирают в детском возрасте. Впрочем, многие так называемые гомозиготы в сушности являются гетерозиготами-компаундами, несушими две различные мутации по LDL-рецепторам.
Тип мутации определяет степень тяжести заболевания, при полном отсутствии рецепторов LDL симптомы более тяжелые, чем при снижении числа рецепторов. Истинным гомозиготам медикаментозное лечение не помогает, в этих случаях необходимы другие подходы, например шунт портальной вены. В одном случае, чтобы обеспечить синтез нормальных рецепторов LDL, больному была сделана трансплантация печени.
Действительно, уровень LDL-холестерина у больного снизился. [c.124]
Окислительное декарбоксилирование ПВК является одной из ключевых реакции в обмене углеводов. В результате этой реакции ПВК, образовавшаяся при окислении глюкозы, включается в главный метаболический путь клетки цикл Кребса, где окисляется до углекислоты и воды с выделением энергии.
Таким образом, благодаря реакции окислительного декарбоксилирования ПВК создаются условия для полного окисления углеводов и утилизации всей заключенной в них энергии Кроме того, образующаяся при действии ПДГ-комплек-са активная форма уксусной кислоты служит источником для синтеза многих биологических продуктов жирных кислот, холестерина, стероидных гормонов, ацетоновых тел и других. [c.16]
Источник: https://www.chem21.info/info/144092/